Суммарные уравнения и частные реакции фотосинтеза. Фотосинтез. Общее уравнение фотосинтеза. Суточный ход фотосинтеза
трансформации
поглощенной организмом
энергии света в
химическую энергию
органических
(неорганических)
соединений.
Главная роль восстановление СО2 до
уровня углеводов с
использованием энергии
света.
Развитие учения о фотосинтезе
Климе́нт Арка́дьевич Тимиря́зев(22 мая (3 июня) 1843, Петербург- 28
апреля 1920, Москва) Научные труды
Тимирязева, посвящены вопросу о
разложении атмосферной углекислоты
зелёными растениями под влиянием
солнечной энергии. Изучение состава и
оптических свойств зелёного пигмента
растений (хлорофилла), его генезиса,
физических и химических условий
разложения углекислоты, определение
составных частей солнечного луча,
принимающих участие в этом явлении,
изучение количественного отношения
между поглощенной энергией и
произведённой работой.Джозеф Пристли (13 марта
1733-6 февраля 1804) -
британский священникдиссентер, естествоиспытатель,
философ, общественный деятель.
Вошёл в историю прежде всего
как выдающийся химик,
открывший кислород и
углекислый газПьер Жозеф Пельтье - (22 марта 1788 - 19 июля
1842) - французский химик и фармацевт, один из
основателей химии алкалоидов.
В 1817 году, вместе с Жозеф Бьенеме Каванту, он
выделил зелёный пигмент из листьев растений, который
они назвали хлорофиллом.Алексей Николаевич Бах
(5 (17) марта 1857 - 13 мая,
1946) - советский биохимик и
физиолог растений. Высказал
мысль о том, что ассимиляция СО2
при фотосинтезе является
сопряженным окислительновосстановительным процессом,
происходящим за счет водорода и
гидроксила воды, причем кислород
выделяется из воды через
промежуточные перекисные
соединения.
Общее уравнение фотосинтеза
6 СО2 + 12 Н2ОС6Н12О6 + 6 О2 + 6 Н2ОУ высших растений фотосинтез осуществляется в
специализированных клетках органоидов листьев –
хлоропластах.
Хлоропласты – это округлые, или дискообразные
тельца длиной 1-10 мкм, толщиной до 3 мкм. Содержание
их в клетках от 20 до 100.
Химический состав (% на сухую массу):
Белок - 35-55
Липиды – 20-30
Углеводы – 10
РНК – 2-3
ДНК – до 0,5
Хлорофилл – 9
Каротиноиды – 4,5
Строение Хлоропласта
10. Происхождение хлоропластов
Виды формирования хлоропластов:Деление
Почкование
Ядерный путь
темнота
ядро
инициальная
частица
свет
проламиллярное
тело
пропластида
хлоропласт
схема ядерного пути
11. Онтогенез хлоропластов
12.
Хлоропласты - зелёные пластиды, которыевстречаются в клетках растений и водорослей.
Ультраструктура хлоропласта:
1. наружняя мембрана
2. межмембранное
пространство
3. внутренняя мембрана
(1+2+3: оболочка)
4. строма (жидкость)
5. тилакоид с просветом
6. мембрана тилакоида
7. грана (стопка тилакоидов)
8. тилакоид (ламела)
9. зерно крахмала
10. рибосома
11. пластидная ДНК
12. плстоглобула (капля жира)
13. Пигменты фотосинтезирующих растений
хлорофиллыфикобилины
Фикобилины
каротиноиды
флавоноидные
пигменты
14. Хлорофиллы
Хлорофи́лл -зелёный пигмент,
обусловливающий
окраску хлоропластов
растений в зелёный
цвет. По химическому
строению
хлорофиллы -
магниевые комплексы
различных
тетрапирролов.
Хлорофиллы имеют
порфириновое
строение.
15.
ХлорофиллыХлорофилл «а»
(сине-зеленые
бактерии)
Хлорофилл «c»
(бурые водоросли)
Хлорофилл «b»
(высшие растения,
зеленые, харовые
водоросли)
Хлорофилл «d»
(красные водоросли)
16. Фикобилины
Фикобилины – этопигменты,
представляющие собой
вспомогательные
фотосинтетические
пигменты, которые могут
передавать энергию
поглощенных квантов
света на хлорофилл,
расширяя спектр действия
фотосинтеза.
Открытые тетрапиррольные
структуры.
Встречаются у водорослей.
17. Каротиноиды
Структурная формула18.
Каротиноиды – этожирорастворимые
пигменты желтого,
красного и оранжевого
цвета. Придают
окраску большинству
оранжевых овощей и
фруктов.
19. Группы каротиноидов:
Каротины - жёлтооранжевый пигмент,непредельный углеводород
из группы каротиноидов.
Формула С40H56. Нерастворим
в воде, но растворяется в
органических растворителях.
Содержится в листьях всех растений, а также в
корне моркови, плодах шиповника и др. Является
провитамином витамина А.
2.
Ксантофиллы - растительный пигмент,
кристаллизуется в призматических кристаллах
жёлтого цвета.
1.
20. Флавоноидные пигменты
Флавоноиды -это группаводорастворимых природных
фенольных соединений.
Представляют собой
гетероциклические
кислородсодержащие
соединения преимущественно
желтого, оранжевого, красного
цвета. Они принадлежат к
соединениям С6-С3-С6 ряда -
в их молекулах имеются два
бензольных ядра, соединенных
друг с другом трехуглеродным
фрагментом.
Структура флавонов
21. Флавоноидные пигменты:
Антоцианы - природные вещества, красящие растения;относятся к гликозидам.
Флавоны и флавонолы. Играют роль поглотителей УФлучей тем самым предохраняют хлорофилл и цитоплазму
от разрушения.
22. Стадии фотосинтеза
световаяОсуществляется в
гранах хлоропластов.
Протекает при наличии
света Быстрые < 10 (-5)
сек
темновая
Осуществляется в
бесцветной белковой строме
хлоропластов.
Для протекания свет
необязателен
Медленные ~ 10 (-2) сек
23.
24.
25. Световая стадия фотосинтеза
В ходе световой стадии фотосинтеза образуютсявысокоэнергетические продукты: АТФ, служащий в
клетке источником энергии, и НАДФН, использующийся
как восстановитель. В качестве побочного продукта
выделяется кислород.
Общее уравнение:
АДФ + Н3РО4 + Н2О + НАДФ
АТФ + НАДФН + 1/2О2
26.
Спектры поглощенияФАР: 380 – 710 нм
Каротиноиды: 400550 нм главный
максимум: 480 нм
Хлорофиллы:
в красной области спектра
640-700 нм
в синей - 400-450 нм
27. Уровни возбуждения хлорофилла
1 уровень. Связан с переходом на более высокийэнергетический уровень электронов в системе
сопряжения двух связей
2 уровень. Связан с возбуждением неспаренных электронов
четырех атомов азота и кислорода в порфириновом
кольце.
28. Пигментные системы
Фотосистема IСостоит из 200 молекул
хлорофилла «а»,50
молекул кароиноидов и 1
молекулы пигмента
(Р700)
Фотосистема II
Состоит из 200 молекул
хлорофилла «а670», 200
молекул хлорофилла «b» и
одной молекулы пигмента
(Р680)
29. Локализация электрон и протон транспортных реакций в тилакоидной мембране
30. Нециклическое фотосинтетическое фосфорилирование (Z – схема, или схема Говинджи)
xе
Фg е
Фф е
НАДФ
Пх
е
FeS
е
АДФ
Цит b6
е
II ФС
НАДФН
АТФ
е
I ФС
Цит f
е
е
Пц
е
Р680
hV
О2
е
Н2 О
Р700
hV
Фф – феофетин
Пx – пластохинон
FeS – железосерный белок
Цит b6 – цитохром
Пц – пластоционин
Фg – феродоксин
х – неизвестное прир.
соединение
31. Фотосинтетическое фосфорилирование
Фотосинтетическое фосфорилирование – это процессобразования энергии АТФ и НАДФН при фотосинтезе с
использованием квантов света.
Виды:
нециклическое (Z-схема).Принимают участие две
пигментные системы.
циклическое. Принимает участие фотосистема I.
псевдоциклическое. Идет по типу нециклического, но не
наблюдается видимого выделения кислорода.
32. Циклическое фотосинтетическое фосфорилирование
еАДФ
Фg
е
АТФ
Цитb6
е
e
Цитf
е
P700
hV
е
АДФ
АТФ
Цит b6 – цитохром
Фg – феродоксин
33. Циклический и нециклический транспорт электронов в хлоропластах
34.
Химизм фотосинтезаФотосинтез
осуществляется
путем
последовательного чередования двух фаз:
световой,
протекающей
с
большой
скоростью и не зависящей от температуры;
темновой, названной так потому, что для
происходящих в этой фазе реакций
световая энергия не требуется.
35. Темновая стадия фотосинтеза
В темновой стадии с участием АТФ и НАДФНпроисходит восстановление CO2 до глюкозы (C6H12O6).
Хотя свет не требуется для осуществления данного
процесса, он участвует в его регуляции.
36. С3-фотосинтез, цикл Кальвина
Цикл Кальвина или восстановительныйпентозофосфатный цикл состоит из трёх стадий:
Карбоксилирования РДФ.
Восстановления. Происходит восстановление 3-ФГК до
3-ФГА.
Регенерация акцептора РДФ. Осуществляются в серии
реакций взаимопревращений фосфорилируемых сахаров с
различным числом углеродных атомов (триоз, тетроз,
пентоз, гексоз, и т.д.)
37. Общее уравнение цикла Кальвина
Н2СО (Р)С=О
НО-С-Н + * СО2
Н-С-ОН
Н2СО (Р)
РДФ
Н2*СО (Р)
2 НСОН
СООН
3-ФГК
Н2*СО (Р)
2НСОН
СОО (Р)
1,3-ФГК
Н2*СО (Р)
2НСОН
С=О
Н
3-ФГА
Н2*СО (Р)
2С=О
НСОН
3-ФДА
конденсация, или
полимеризация
Н
Н2СО (Р)
Н2СО (Р)
С=О
С=О
С=О
НСОН
НОСН
НОСН
НОСН
Н*СОН
НСОН
Н*СОН
НСОН
НСОН
НСОН
Н2СО (Р)
Н2СОН
Н2СО (Р)
1,6-дифосфат- фруктозо-6глюкоза-6фруктоза
фосфат
фосфат
Н
С=О
НСОН
НОСН
Н*СОН
НСОН
Н2СОН
глюкоза
38. С4-фотосинтез (путь Хэтча – Слэка – Карпилова)
Осуществляется у растений с двумя типами хлоропласта.Акцептором СО2 помимо РДФ может быть трех
углеродное соединение – фосфоэнол ПВК (ФЕП)
C4 –путь был впервые обнаружен
у тропических злаков. В работах
Ю.С.Карпилова, М.Хэтча, К.Слэка с
использованием меченого углерода
было показано, что первыми
продуктами фотосинтеза у этих
растений являются органические
кислоты.
39.
40. Фотосинтез по типу толстянковых
Характерно для растенийсуккуленотов.В ночное время
фиксируют углерод в
органические кислоты по
преимуществу в яблочные. Это
происходит под действием
ферментов
пируваткарбокислазы. Это
позволяет в течении дня
держать устьица закрытыми и
таким образом сокращать
транспирацию. Этот тип
получил название САМфотосинтез.
41. САМ фотосинтез
При CAM фотосинтезе происходит разделениеассимиляции CO2 и цикла Кальвина не в
пространстве как у С4, а во времени. Ночью в
вакуолях клеток по аналогичному
вышеописанному механизму при открытых
устьицах накапливается малат, днём при
закрытых устьицах идёт цикл Кальвина. Этот
механизм позволяет максимально экономить
воду, однако уступает в эффективности и С4, и
С3.
42.
43.
Фотодыхание44. Влияние внутренних и внешних факторов на фотосинтез
Фотосинтеззначительно
изменяется из-за
влияния на него
комплекса часто
взаимодействующих
внешних и внутренних
факторов.
45. Факторы, влияющие на фотосинтез
1.Онтогенетическое
состояние растения.
Максимальная
интенсивность
фотосинтеза наблюдается
во время перехода
растений от вегетации в
репродуктивную фазу. У
стареющих листьев
интенсивность
фотосинтеза значительно
падает.
46. Факторы, влияющие на фотосинтез
2. Свет. В темноте фотосинтез не происходит, так какобразующийся при дыхании углекислый газ выделяется из
листьев; с увеличением интенсивности света достигается
компенсационная точка при которой поглощение
углекислого газа при фотосинтезе и ее освобождение при
дыхании уравновешивают друг друга.
47. Факторы, влияющие на фотосинтез
3. Спектральныйсостав света.
Спектральный
состав солнечного
света испытывает
некоторые
изменения в
течении суток и в
течении года.
48. Факторы, влияющие на фотосинтез
4. СО2.Является основным
субстратом фотосинтеза и от
его содержания зависит
интенсивность этого процесса.
В атмосфере содержится
0,03% по объему; увеличение
объема углекислого газа от 0,1
до 0,4% увеличивает
интенсивность фотосинтеза до
определенного предела, а
затем сменяется
углекислотным насыщением.
49. Факторы, влияющие на фотосинтез
5.Температура.У растений умеренной
зоны оптимальная
температура для
фотосинтеза
является 20-25; у
тропических – 2035.
50. Факторы, влияющие на фотосинтез
6. Содержание воды.Снижение обезвоженности тканей более чем на 20%
приводит к уменьшению интенсивности фотосинтеза и к
его дальнейшему прекращению, если потеря воды будет
более 50%.
51. Факторы, влияющие на фотосинтез
7. Микроэлементы.Недостаток Fe
вызывает хлороз и
влияет на активность
ферментов. Mn
необходим для
освобождения
кислорода и для
усвоения углекислого
газа. Недостаток Cu и
Zn снижает фотосинтез
на 30%
52. Факторы, влияющие на фотосинтез
8.Загрязняющиевещества и
химические
препараты.
Вызывают
снижение
фотосинтеза.
Наиболее
опасные
вещества: NO2,
SO2, взвешенные
частицы.
53. Суточный ход фотосинтеза
При умеренной дневной температуре и достаточнойвлажности дневной ход фотосинтеза примерно
соответствует изменению интенсивности солнечной
инсоляции. Фотосинтез, начинаясь утром с восходом
солнца, достигает максимума в полуденные часы,
постепенно снижается к вечеру и прекращается с заходом
солнца. При повышенной температуре и уменьшении
влажности максимум фотосинтеза сдвигается на ранние
часы.
54. Вывод
Таким образом фотосинтез – единственный процесс наЗемле, идущий в грандиозных масштабах, связанный с
превращением энергии солнечного света в энергию химических
связей. Эта энергия, запасенная зелеными растениями,
составляет основу для жизнедеятельности всех других
гетеротрофных организмов на Земле от бактерий до человека.
Фотосинтетическое фосфорилирование было обнаружено Д. Арноном с сотрудниками и другими исследователями в опытах с изолированными хлоропластами высших растений и с бесклеточными препаратами из различных фотосинтезирующих бактерий и водорослей. При фотосинтезе происходят два типа фотоосинтетического фосфорилирования: циклическое и нециклическое. При обоих видах фотофосфорилирования синтез АТР из ADP и неорганического фосфата происходит на этапе передачи электронов от цитохрома b6 к цитохрому f.
Синтез АТР осуществляется при участии АТР-азного комплекса, «вмонтированного» в белково-липидную мембрану тилакоида с eе внешней стороны. Согласно теории Митчелла, так же, как и в случае окислительного фосфорилирования в митохондрии, находящаяся в мембране тилакоида цепь переноса электронов функционирует как «протонный насос», создавая градиент концентрации протонов. Однако в данном случае происходящий при поглощении света перенос электронов вызывает их перемещение снаружи во внутрь тилакоида и возникающий трансмембранный потенциал (между внутренней и внешней поверхностью мембраны) обратен тому, который образуется в мембране митохондрии. Электростатическая энергия и энергия градиента протонов используется для синтеза АТР ATP-синтетазой.
При нециклическом фотофосфорилировании электроны, поступившие от воды и соединения Z к фотосистеме 2, а затем к фотосистеме 1, направляются к промежуточному соединению Х, а затем используются на восстановление NADP+ до NADPH; их путь здесь заканчивается. При циклическом фотофосфорилировании электроны, поступившие от фотосистемы 1 к соединению Х, направляются снова к цитохрому b6 и от него далее к цитохрому У, участвуя на этом последнем этапе своего пути в синтезе АТР из ADP и неорганического фосфата. Таким образом, при нециклическом фотофосфорилировании перемещение электронов сопровождается синтезом АТР и NADPH. При циклическом фотофосфорилировании происходит только синтез АТР, а NADPH не образуется. АТР, образовавшийся в процессе фотофосфорилирования и дыхания, используется не только при восстановлении фосфоглицериновой кислоты до углевода, но и в других синтетических реакциях - при синтезе крахмала, белков, липидов, нуклеиновых кислот и пигментов. Он также служит источником энергии для процессов движения, транспорта метаболитов, поддержания ионного баланса и т.д.
Роль пластохинонов в фотосинтезе
В хлоропластах открыты пять форм пластохинонов, обозначаемых буквами А, В, С, D и Е, являющихся производными бензохинона. Так, например, пластохинон А представляет собой 2,З-диметил-5-соланезилбензохинон. Пластохиноны весьма близки по строению к убихинонам (коэнзимам Q), играющим важную роль в процессе переноса электронов при дыхании. Важная роль пластохинонов в процессе фотосинтеза следует из того факта, что если их экстрагировать из хлоропластов петролейным эфиром, то фотолиз воды и фотофосфорилирование прекращаются, но возобновляются после добавки пластохинонов. Каковы детали функциональной взаимосвязи различных пигментов и переносчиков электронов, участвующих в процессе фотосинтеза, - цитохромов, ферредоксина, пластоцианина и пластохинонов, - должны показать дальнейшие исследования. Во всяком случае, каковы бы ни были детали этого процесса, в настоящее время очевидно, что световая фаза фотосинтеза приводит к образованию трех специфических продуктов: NADPH, АТР и молекулярного кислорода.
Какие соединения образуются в результате третьего, темнового этапа фотосинтеза?
Существенные результаты, проливающие свет на природу первичных продуктов, образующихся при фотосинтезе, получены с помощью изотопной методики. В этих исследованиях растения ячменя, а также одноклеточные зеленые водоросли Chlorella и Scenedesmus получали в качестве источника углерода углекислый газ, содержавший меченый радиоактивный углерод 14C. После чрезвычайно кратковременного облучения подопытных растений, исключавшего возможность вторичных реакций, исследовалось распределение изотопного углерода в различных продуктах фотосинтеза. Было установлено, что первый продукт фотосинтеза - фосфоглицериновая кислота; вместе с тем при весьма кратковременном облучении растений наряду с фосфоглицериновой кислотой образуется незначительное количество фосфоенолпировиноградной и яблочной кислот. Например, в опытах с одноклеточной зеленой водорослью Sceriedesmus после фотосинтеза, продолжавшегося пять секунд, 87% изотопного углерода было обнаружено в составе фосфоглицериновой кислоты, 10% - в фосфоенолпировиноградной кислоте и 3% -- в яблочной кислоте. По-видимому, фосфоенолпировиноградная кислота является продуктом вторичного превращения фосфоглицериновой кислоты. При более длительном фотосинтезе, продолжающемся 15-60 секунд, радиоактивный углерод 14C обнаруживается также в гликолевой кислоте, триозофосфатах, сахарозе, аспарагиновой кислоте, аланине, серине, гликоколе, а также в белках. Позже всего меченый углерод обнаруживается в глюкозе, фруктозе, янтарной, фумаровой и лимонной кислотах, а также в некоторых аминокислотах и амидах (треонин, фенилаланин, тирозин, глютамин, аспарагин). Таким образом, опыты с усвоением растениями углекислого газа, содержащего меченый углерод, показали, что первым продуктом фотосинтеза является фосфоглицериновая кислота.
К какому веществу присоединяется углекислый газ в процессе фотосинтеза?
Работы М. Кальвина, проведенные с помощью радиоактивного углерода 14С, показали, что у большинства растений соединением, к которому присоединяется СО2, является рибулозодифосфат. Присоединяя СО2, он дает две молекулы фосфоглицериновой кислоты. Последняя фосфоорилируется при участии АТР с образованием дифосфоглицериновой кислоты, которая при участии NADPH восстанавливается и образует фосфоглицериновый альдегид, частично превращающийся в фосфодиоксиацетон. Благодаря синтетическому действию фермента альдолазы, фосфоглицериновый альдегид и фосфодиоксиацетон, соединяясь, образуют молекулу фруктозодифосфата, из которого далее синтезируются сахароза и различные полисахариды. Рибулозодифосфат - акцептор СО2, образуется в результате ряда ферментативных превращений фосфоглицеринового альдегида, фосфодиоксиацетона и фруктозодифосфата. В качестве промежуточных продуктов при этом возникают эритрозофосфат, седогептулозофосфат, ксилулозофосфат, рибозофосфат и рибулозофосфат. Ферментные системы, катализирующие все эти превращения, найдены в клетках хлореллы, в листьях шпината и в других растениях. Согласно М. Кальвину, процесс образования фосфоглицериновой кислоты из рибулозодифосфата и СО2 носит циклический характер. Ассимиляция углекислого газа с образованием фосфоглицериновой кислоты происходит без участия света и хлорофилла и является темновым процессом. Водород воды в конечном счете используется на восстановление фосфоглицериновой кислоты до фосфоглицеринового альдегида. Этот процесс катализируется ферментом дегидрогеназой фосфоглицеринового альдегида и в качестве источника водорода требует участия NADPH. Так как этот процесс в темноте немедленно прекращается, очевидно, что восстановление NADP осуществляется водородом, образующимся при фотолизе воды.
Уравнение Кальвина для фотосинтеза
Суммарное уравнение цикла Кальвина имеет следующий вид:
6СО2 + 12NADPH + 12Н+ + 18АТР + 11Н2О = фруктозо-б-фосфат + 12NADP+ + 18ADP + 17Р неорг
Таким образом, для синтеза одной молекулы гексозы требуется шесть молекул СО2. Для превращения одной молекулы СО2 нужно две молекулы NADPH и три молекулы АТР (1: 1,5). Так как при нециклическом фотофосфорилировании отношение образующихся NADPH:АТР составляет 1:1, добавочное необходимое количество АТР синтезируется в процессе циклического фотофосфорилирования.
Путь углерода при фотосинтезе изучался Кальвином при сравнительно высоких концентрациях СО2. При более низких концентрациях, приближающихся к атмосферным (0,03%), в хлоропласте под действием рибулозодифосфаткарбоксилазы образуется значительное количество фосфогликолевой кислоты. Последняя в процессе транспорта через мембрану хлоропласта гидролизуется специфической фосфатазой, и образовавшаяся гликолевая кислота перемещается из хлоропласта в связанные с ним субклеточные структуры - пероксисомы, где под действием фермента гликолатоксидазы окисляется до глиоксилевой кислоты HOC-COOH. Последняя путем переаминирования образует глицин, который, перемещаясь в митохондрию, превращается здесь в серин.
Это превращение сопровождается образованием СО2 и NН3: 2 глицин + Н2О = серин + СО2 + NН3 +2Н+ +2е-.
Однако аммиак не выделяется во внешнюю среду, а связывается в виде глютамина. Таким образом, пероксисомы и митохондрии принимают участие в процессе так называемого фотодыхания - стимулируемого светом процесса поглощения кислорода и выделения СО2. Этот процесс связан с превращениями гликолевой кислоты и ее окислением до СО2. В результате интенсивного фотодыхания может значительно (до 30%) снижаться продуктивность растений.
Другие возможности усвоения СО2 в процессе фотосинтеза
Усвоение СО2 в процессе фотосинтеза происходит не только путем карбоксилирования рибулозодифосфата, но и путем карбоксилирования других соединений. Например, показано, что у сахарного тростника, кукурузы, сорго, проса и ряда других растений особенно важную роль в процессе фотосинтетической фиксации играет фермент фосфоенолпируват-карбоксилаза, синтезирующая из фосфоенолпирувата, СО2 и воды щавелевоуксусную кислоту. Растения, у которых первым продуктом фиксации СО2 является фосфоглицериновая кислота, принято называть С3-растениями, а те, у которых синтезируется щавелевоуксусная кислота, -C4-растениями. Упоминавшийся выше процесс фотодыхания характерен для С3-растений и является следствием ингибирующего действия кислорода на рибулозодифосфат-карбоксилазу.
Фотосинтез у бактерий
У фотосинтезирующих бактерий фиксация СО2 происходит при участии ферредоксина. Так, из фотосинтезирующей бактерии Chromatium выделена и частично очищена ферментная система, которая при участии ферредоксина катализирует восстановительный синтез пировиноградной кислоты из СО2 и ацетилкоэнзима А:
Ацетил-СоА + С02 + ферредоксин восстановл. = пируват + ферредоксин окислен. + СоА
Аналогичным образом при участии ферредоксина в бесклеточных ферментных препаратах, выделенных из фотосинтезирующей бактерии Chlorobium thiosulfatophilum , происходит синтез а-кетоглютаровой кислоты путем карбоксилирования янтарной кислоты:
Сукцинил-СоА + СО2 + ферредоксин восстановл. = a-кетоглютарат + СоА + ферредоксин окислен.
У некоторых микроорганизмов, содержащих бактериохлорофилл, так называемых пурпурных серобактерий, на свету также происходит процесс фотосинтеза. Однако в отличие от фотосинтеза высших растений в данном случае восстановление углекислого газа осуществляется сероводородом. Суммарное уравнение фотосинтеза у пурпурных бактерий можно представить следующим образом:
Свет, бактериохлорофилл: CO2 + 2H2S = CH2O + H2O + 2S
Таким образом, и в данном случае фотосинтез представляет собой сопряженный окислительно-восстановительный процесс, идущий под влиянием поглощенной бактериохлорофиллом световой энергии. Из приведенного уравнения видно, что в результате фотосинтеза пурпурные бактерии выделяют свободную серу, которая накапливается в них в виде гранул.
Исследования, проведенные при помощи изотопной методики с анаэробной фотосинтезирующей пурпурной бактерией Chromatium, показали, что при очень коротких сроках фотосинтеза (30 секунд) около 45% углерода СО2 включается в аспарагиновую кислоту, а около 28% - в фосфоглицериновую кислоту. По-видимому, образование фосфоглицериновой кислоты предшествует образованию аспарагиновой кислоты, а наиболее ранним продуктом фотосинтеза у Chromatium, так же как у высших растений и одноклеточных зеленых водорослей, является рибулозодифосфат. Последний под действием рибулозодифосфаткарбоксилазы присоединяет СО2 с образованием фосфоглицериновой кислоты. Эта кислота у Chromatium в соответствии со схемой Кальвина может частично превращаться в фосфорилированные сахара, а в основном превращается в аспарагиновую кислоту. Образование аспарагиновой кислоты происходит путем превращения фосфоглицериновой кислоты в фосфоенолпировиноградную кислоту, которая, подвергаясь карбоксилированию, дает щавелевоуксусную кислоту; последняя путем переаминирования дает аспарагиновую кислоту.
Фотосинтез - источник органических веществ на Земле
Процесс фотосинтеза, происходящий при участии хлорофилла, в настоящее время - главный источник образования органического вещества на Земле.
Фотосинтез для получения водорода
Нужно отметить, что одноклеточные фотосинтезирующие водоросли в анаэробных условиях выделяют газообразный водород. Изолированные хлоропласты высших растений, освещаемые в присутствии фермента гидрогеназы катализирующего реакцию 2Н+ + 2е- = Н2, также выделяют водород. Таким образом, возможно фотосинтетическое получение водорода в качестве топлива. Этот вопрос, особеннo в условиях энергетического кризиса, привлекает к себе большое внимание.
Новый вид фотосинтеза
В. Стокениусом был открыт принципиально новый вид фотосинтеза. Оказалось, что у бактерии Halobacterium halobium , живущей в концентрированных растворах хлористого натрия, в окружающей протоплазму белково-липидной мембране содержится хромопротеид бактериородопсин, аналогичный родопсину - зрительному пурпуру глаза животных. В бактериородопсине ретиналь (альдегидная форма витамина А) связан с белком, имеющим молекулярную массу равную 26534, он состоит из 247 аминокислотных остатков. Поглощая свет, бактериородопсин участвует в процессе превращения световой энергии в химическую энергию высокоэнергетических связей АТР. Таким образом, не содержащий хлорофилла организм способен с помощью бактериородопсина использовать световую энергию для синтеза АТР и обеспечения клетки энергией.
Уравнение: 6СО2 + 6Н2О ----> С6Н12О6 + 6О2
Фотосинтез - процесс образования органического вещества из углекислого газа и воды на свету при участии фотосинтетических пигментов (хлорофилл у растений, бактериохлорофилл и бактериородопсин у бактерий).
Фотосинтез является основным источником биологической энергии, фотосинтезирующие автотрофы используют её для синтеза органических веществ из неорганических, гетеротрофы существуют за счёт энергии, запасённой автотрофами в виде химических связей, высвобождая её в процессах дыхания и брожения. Энергия получаемая человечеством при сжигании ископаемого топлива (уголь, нефть, природный газ, торф) также является запасённой в процессе фотосинтеза.
Фотосинтез является главным входом неорганического углерода в биологический цикл. Весь свободный кислород атмосферы - биогенного происхождения и является побочным продуктом фотосинтеза. Формирование окислительной атмосферы (кислородная катастрофа) полностью изменило состояние земной поверхности, сделало возможным появление дыхания, а в дальнейшем, после образования озонового слоя, позволило жизни выйти на сушу.
Бактериальный фотосинтез
Некоторые пигментосодержащие серобактерии (пурпурные, зелёные), содержащие специфические пигменты – бактериохлорофиллы, способны поглощать солнечную энергию, с помощью которой сероводород в их организмах расщепляется и отдаёт атомы водорода для восстановления соответствующих соединений. Этот процесс имеет много общего с фотосинтезом и отличается только тем, что у пурпурных и зелёных бактерий донором водорода является сероводород (изредка – карбоновые кислоты), а у зелёных растений – вода. У тех и других отщепление и перенесение водорода осуществляется благодаря энергии поглощённых солнечных лучей.
Такой бактериальный фотосинтез, который происходит без выделения кислорода, называется фоторедукцией. Фоторедукция углекислого газа связана с перенесением водорода не от воды, а от сероводорода:
6СО 2 +12Н 2 S+hv → С6Н 12 О 6 +12S=6Н 2 О
Биологическое значение хемосинтеза и бактериального фотосинтеза в масштабах планеты относительно невелико. Только хемосинтезирующие бактерии играют существенную роль в процессе круговорота серы в природе. Поглощаясь зелёными растениями в форме солей серной кислоты, сера восстанавливается и входит в состав белковых молекул. Далее при разрушении отмерших растительных и животных остатков гнилостными бактериями сера выделяется в виде сероводорода, который окисляется серобактериями до свободной серы (или серной кислоты), образующий в почве доступные для растения сульфиты. Хемо- и фотоавтотрофные бактерии имеют существенное значение в круговороте азота и серы.
Фотосинтез - это преобразование энергии света в энергию химических связей органических соединений.
Фотосинтез характерен для растений, в том числе всех водорослей, ряда прокариот, в том числе цианобактерий, некоторых одноклеточных эукариот.
В большинстве случаев при фотосинтезе в качестве побочного продукта образуется кислород (O 2). Однако это не всегда так, поскольку существует несколько разных путей фотосинтеза. В случае выделения кислорода его источником является вода, от которой на нужды фотосинтеза отщепляются атомы водорода.
Фотосинтез состоит из множества реакций, в которых участвуют различные пигменты, ферменты, коферменты и др. Основными пигментами являются хлорофиллы, кроме них - каротиноиды и фикобилины.
В природе распространены два пути фотосинтеза растений: C 3 и С 4 . У других организмов есть своя специфика реакций. Все, что объединяет эти разные процессы под термином «фотосинтез», – во всех них в общей сложности происходит преобразование энергии фотонов в химическую связь. Для сравнения: при хемосинтезе происходит преобразование энергии химической связи одних соединений (неорганических) в другие - органические.
Выделяют две фазы фотосинтеза - световую и темновую. Первая зависит от светового излучения (hν), которое необходимо для протекания реакций. Темновая фаза является светонезависимой.
У растений фотосинтез протекает в хлоропластах. В результате всех реакций образуются первичные органические вещества, из которых потом синтезируются углеводы, аминокислоты, жирные кислоты и др. Обычно суммарную реакцию фотосинтеза пишут в отношении глюкозы - наиболее распространенного продукта фотосинтеза :
6CO 2 + 6H 2 O → C 6 H 12 O 6 + 6O 2
Атомы кислорода, входящие в молекулу O 2 , берутся не из углекислого газа, а из воды. Углекислый газ – источник углерода , что более важно. Благодаря его связыванию у растений появляется возможность синтеза органики.
Представленная выше химическая реакция есть обобщенная и суммарная. Она далека от сути процесса. Так глюкоза не образуется из шести отдельных молекул углекислоты. Связывание CO 2 происходит по одной молекуле, которая сначала присоединяется к уже существующему пятиуглеродному сахару.
Для прокариот характерны свои особенности фотосинтеза. Так у бактерий главный пигмент - бактериохлорофилл, и не выделяется кислород, так как водород берется не из воды, а часто из сероводорода или других веществ. У сине-зеленых водорослей основным пигментом является хлорофилл, и при фотосинтезе выделяется кислород.
Световая фаза фотосинтеза
В световой фазе фотосинтеза происходит синтез АТФ и НАДФ·H 2 за счет лучистой энергии. Это происходит на тилакоидах хлоропластов , где пигменты и ферменты образуют сложные комплексы для функционирования электрохимических цепей, по которым передаются электроны и отчасти протоны водорода.
Электроны в конечном итоге оказываются у кофермента НАДФ, который, заряжаясь отрицательно, притягивает к себе часть протонов и превращается в НАДФ·H 2 . Также накопление протонов по одну сторону тилакоидной мембраны и электронов по другую создает электрохимический градиент, потенциал которого используется ферментом АТФ-синтетазой для синтеза АТФ из АДФ и фосфорной кислоты.
Главными пигментами фотосинтеза являются различные хлорофиллы. Их молекулы улавливают излучение определенных, отчасти разных спектров света. При этом некоторые электроны молекул хлорофилла переходят на более высокий энергетический уровень. Это неустойчивое состояние, и по-идее электроны путем того же излучения должны отдать в пространство полученную из вне энергию и вернуться на прежний уровень. Однако в фотосинтезирующих клетках возбужденные электроны захватываются акцепторами и с постепенным уменьшением своей энергии передаются по цепи переносчиков.
На мембранах тилакоидов существуют два типа фотосистем, испускающих электроны при действия света. Фотосистемы представляют собой сложный комплекс большей частью хлорофильных пигментов с реакционным центром, от которого и отрываются электроны. В фотосистеме солнечный свет ловит множество молекул, но вся энергия собирается в реакционном центре.
Электроны фотосистемы I, пройдя по цепи переносчиков, восстанавливают НАДФ.
Энергия электронов, оторвавшихся от фотосистемы II, используется для синтеза АТФ. А сами электроны фотосистемы II заполняют электронные дырки фотосистемы I.
Дырки второй фотосистемы заполняются электронами, образующимися в результате фотолиза воды . Фотолиз также происходит при участии света и заключается в разложении H 2 O на протоны, электроны и кислород. Именно в результате фотолиза воды образуется свободный кислород. Протоны участвуют в создании электрохимического градиента и восстановлении НАДФ. Электроны получает хлорофилл фотосистемы II.
Примерное суммарное уравнение световой фазы фотосинтеза:
H 2 O + НАДФ + 2АДФ + 2Ф → ½O 2 + НАДФ · H 2 + 2АТФ
Циклический транспорт электронов
Выше описана так называемый нецикличная световая фаза фотосинтеза . Есть еще циклический транспорт электронов, когда восстановления НАДФ не происходит . При этом электроны от фотосистемы I уходят на цепь переносчиков, где идет синтез АТФ. То есть эта электрон-транспортная цепь получает электроны из фотосистемы I, а не II. Первая фотосистема как бы реализует цикл: в нее возвращаются ей же испускаемые электроны. По дороге они тратят часть своей энергии на синтез АТФ.
Фотофосфорилирование и окислительное фосфорилирование
Световую фазу фотосинтеза можно сравнить с этапом клеточного дыхания - окислительным фосфорилированием, которое протекает на кристах митохондрий. Там тоже происходит синтез АТФ за счет передачи электронов и протонов по цепи переносчиков. Однако в случае фотосинтеза энергия запасается в АТФ не для нужд клетки, а в основном для потребностей темновой фазы фотосинтеза. И если при дыхании первоначальным источником энергии служат органические вещества, то при фотосинтезе – солнечный свет. Синтез АТФ при фотосинтезе называется фотофосфорилированием , а не окислительным фосфорилированием.
Темновая фаза фотосинтеза
Впервые темновую фазу фотосинтеза подробно изучили Кальвин, Бенсон, Бэссем. Открытый ими цикл реакций в последствии был назван циклом Кальвина, или C 3 -фотосинтезом. У определенных групп растений наблюдается видоизмененный путь фотосинтеза – C 4 , также называемый циклом Хэтча-Слэка.
В темновых реакциях фотосинтеза происходит фиксация CO 2 . Темновая фаза протекает в строме хлоропласта.
Восстановление CO 2 происходит за счет энергии АТФ и восстановительной силы НАДФ·H 2 , образующихся в световых реакциях. Без них фиксации углерода не происходит. Поэтому хотя темновая фаза напрямую не зависит от света, но обычно также протекает на свету.
Цикл Кальвина
Первая реакция темновой фазы – присоединение CO 2 (карбоксилировани е ) к 1,5-рибулезобифосфату (рибулезо-1,5-дифосфат ) – РиБФ . Последний представляет собой дважды фосфорилированную рибозу. Данную реакцию катализирует фермент рибулезо-1,5-дифосфаткарбоксилаза, также называемый рубиско .
В результате карбоксилирования образуется неустойчивое шестиуглеродное соединение, которое в результате гидролиза распадается на две трехуглеродные молекулы фосфоглицериновой кислоты (ФГК) – первый продукт фотосинтеза. ФГК также называют фосфоглицератом.
РиБФ + CO 2 + H 2 O → 2ФГК
ФГК содержит три атома углерода, один из которых входит в состав кислотной карбоксильной группы (-COOH):
Из ФГК образуется трехуглеродный сахар (глицеральдегидфосфат) триозофосфат (ТФ) , включающий уже альдегидную группу (-CHO):
ФГК (3-кислота) → ТФ (3-сахар)
На данную реакцию затрачивается энергия АТФ и восстановительная сила НАДФ · H 2 . ТФ - первый углевод фотосинтеза.
После этого большая часть триозофосфата затрачивается на регенерацию рибулозобифосфата (РиБФ), который снова используется для связывания CO 2 . Регенерация включает в себя ряд идущих с затратой АТФ реакций, в которых участвуют сахарофосфаты с количеством атомов углерода от 3 до 7.
В таком круговороте РиБФ и заключается цикл Кальвина.
Из цикла Кальвина выходит меньшая часть образовавшегося в нем ТФ. В перерасчете на 6 связанных молекул углекислого газа выход составляет 2 молекулы триозофосфата. Суммарная реакция цикла с входными и выходными продуктами:
6CO 2 + 6H 2 O → 2ТФ
При этом в связывании участвую 6 молекул РиБФ и образуется 12 молекул ФГК, которые превращаются в 12 ТФ, из которых 10 молекул остаются в цикле и преобразуются в 6 молекул РиБФ. Поскольку ТФ - это трехуглеродный сахар, а РиБФ - пятиуглеродный, то в отношении атомов углерода имеем: 10 * 3 = 6 * 5. Количество атомов углерода, обеспечивающих цикл не изменяется, весь необходимый РиБФ регенерируется. А шесть вошедших в цикл молекул углекислоты затрачиваются на образование двух выходящих из цикла молекул триозофосфата.
На цикл Кальвина в расчете на 6 связанных молекул CO 2 затрачивается 18 молекул АТФ и 12 молекул НАДФ · H 2 , которые были синтезированы в реакциях световой фазы фотосинтеза.
Расчет ведется на две выходящие из цикла молекулы триозофосфата, так как образующаяся в последствии молекула глюкозы, включает 6 атомов углерода.
Триозофосфат (ТФ) - конечный продукт цикла Кальвина, но его сложно назвать конечным продуктом фотосинтеза, так как он почти не накапливается, а, вступая в реакции с другими веществами, превращается в глюкозу, сахарозу, крахмал, жиры, жирные кислоты, аминокислоты. Кроме ТФ важную роль играет ФГК. Однако подобные реакции происходят не только у фотосинтезирующих организмов. В этом смысле темновая фаза фотосинтеза – это то же самое, что цикл Кальвина.
Из ФГК путем ступенчатого ферментативного катализа образуется шестиуглеродный сахар фруктозо-6-фосфат , который превращается в глюкозу . В растениях глюкоза может полимеризоваться в крахмал и целлюлозу. Синтез углеводов похож на процесс обратный гликолизу.
Фотодыхание
Кислород подавляет фотосинтез. Чем больше O 2 в окружающей среде, тем менее эффективен процесс связывания CO 2 . Дело в том, что фермент рибулозобифосфат-карбоксилаза (рубиско) может реагировать не только с углекислым газом, но и кислородом. В этом случае темновые реакции несколько иные.
Фосфогликолат - это фосфогликолевая кислота. От нее сразу отщепляется фосфатная группа, и она превращается в гликолевую кислоту (гликолат). Для его «утилизации» снова нужен кислород. Поэтому чем больше в атмосфере кислорода, тем больше он будет стимулировать фотодыхание и тем больше растению будет требоваться кислорода, чтобы избавиться от продуктов реакции.
Фотодыхание - это зависимое от света потребление кислорода и выделение углекислого газа. То есть обмен газов происходит как при дыхании, но протекает в хлоропластах и зависит от светового излучения. От света фотодыхание зависит лишь потому, что рибулозобифосфат образуется только при фотосинтезе.
При фотодыхании происходит возврат атомов углерода из гликолата в цикл Кальвина в виде фосфоглицериновой кислоты (фосфоглицерата).
2 Гликолат (С 2) → 2 Глиоксилат (С 2) →2 Глицин (C 2) - CO 2 → Серин (C 3) →Гидроксипируват (C 3) → Глицерат (C 3) → ФГК (C 3)
Как видно, возврат происходит не полный, так как один атом углерода теряется при превращении двух молекул глицина в одну молекулу аминокислоты серина, при этом выделяется углекислый газ.
Кислород необходим на стадиях превращения гликолата в глиоксилат и глицина в серин.
Превращения гликолата в глиоксилат, а затем в глицин происходят в пероксисомах, синтез серина в митохондриях. Серин снова поступает в пероксисомы, где из него сначала получается гидрооксипируват, а затем глицерат. Глицерат уже поступает в хлоропласты, где из него синтезируется ФГК.
Фотодыхание характерно в основном для растений с C 3 -типом фотосинтеза. Его можно считать вредным, так как энергия бесполезно тратится на превращения гликолата в ФГК. Видимо фотодыхание возникло из-за того, что древние растения были не готовы к большому количеству кислорода в атмосфере. Изначально их эволюция шла в атмосфере богатой углекислым газом, и именно он в основном захватывал реакционный центр фермента рубиско.
C 4 -фотосинтез, или цикл Хэтча-Слэка
Если при C 3 -фотосинтезе первым продуктом темновой фазы является фосфоглицериновая кислота, включающая три атома углерода, то при C 4 -пути первыми продуктами являются кислоты, содержащие четыре атома углерода: яблочная, щавелевоуксусная, аспарагиновая.
С 4 -фотосинтез наблюдается у многих тропических растений, например, сахарного тростника, кукурузы.
С 4 -растения эффективнее поглощают оксид углерода, у них почти не выражено фотодыхание.
Растения, в которых темновая фаза фотосинтеза протекает по C 4 -пути, имеют особое строение листа. В нем проводящие пучки окружены двойным слоем клеток. Внутренний слой - обкладка проводящего пучка. Наружный слой - клетки мезофилла. Хлоропласты клеток слоев отличаются друг от друга.
Для мезофильных хлоропласт характерны крупные граны, высокая активность фотосистем, отсутствие фермента РиБФ-карбоксилазы (рубиско) и крахмала. То есть хлоропласты этих клеток адаптированы преимущественно для световой фазы фотосинтеза.
В хлоропластах клеток проводящего пучка граны почти не развиты, зато высока концентрация РиБФ-карбоксилазы. Эти хлоропласты адаптированы для темновой фазы фотосинтеза.
Углекислый газ сначала попадает в клетки мезофилла, связывается с органическими кислотами, в таком виде транспортируется в клетки обкладки, освобождается и далее связывается также, как у C 3 -растений. То есть C 4 -путь дополняет, а не заменяет C 3 .
В мезофилле CO 2 присоединяется к фосфоенолпирувату (ФЕП) с образованием оксалоацетата (кислота), включающего четыре атома углерода:
Реакция происходит при участии фермента ФЕП-карбоксилазы, обладающего более высоким сродством к CO 2 , чем рубиско. К тому же ФЕП-карбоксилаза не взаимодействует с кислородом, а значит не затрачивается на фотодыхание. Таким образом, преимущество C 4 -фотосинтеза заключается в более эффективной фиксации углекислоты, увеличению ее концентрации в клетках обкладки и следовательно более эффективной работе РиБФ-карбоксилазы, которая почти не расходуется на фотодыхание.
Оксалоацетат превращается в 4-х углеродную дикарбоновую кислоту (малат или аспартат), которая транспортируется в хлоропласты клеток обкладки проводящих пучков. Здесь кислота декарбоксилируется (отнятие CO 2), окисляется (отнятие водорода) и превращается в пируват. Водород восстанавливает НАДФ. Пируват возвращается в мезофилл, где из него регенерируется ФЕП с затратой АТФ.
Оторванный CO 2 в хлоропластах клеток обкладки уходит на обычный C 3 -путь темновой фазы фотосинтеза, т. е. в цикл Кальвина.
Фотосинтез по пути Хэтча-Слэка требует больше энергозатрат.
Считается, что C 4 -путь возник в эволюции позже C 3 и во многом является приспособлением против фотодыхания.
1. Дайте определения понятий.
Фотосинтез
- процесс образования органических веществ из углекислого газа и воды на свету при участии фотосинтетических пигментов.
Автотрофы
- организмы, синтезирующие органические вещества из неорганических.
Гетеротрофы - организмы, которые не способны синтезировать органические вещества из неорганических путем фотосинтеза или хемосинтеза.
Миксотрофы
- организмы, способные использовать различные источники углерода и энергии.
2. Заполните таблицу.
3. Заполните таблицу.

4. Объясните суть утверждения великого русского ученого К. А. Тимирязева: «Полено - это консерв солнечной энергии».
Полено – часть дерева, ткани его состоят из накопленных органических соединений (целлюлоза, сахара и др.), которые образовались в процессе фотосинтеза.
5. Напишите суммарное уравнение фотосинтеза. Не забудьте указать обязательные условия протекания реакций.

12. Выберите термин и объясните, насколько его современное значение соответствует первоначальному значению его корней.
Выбранный термин – миксотрофы.
Соответствие. Термин уточнен, так называют организмы со смешанными типом питания, которые способны использовать различные источники углерода и энергии.
13. Сформулируйте и запишите основные идеи § 3.3.
По типу питания все живые организмы делятся на:
Автотрофы, синтезирующие органические вещества из неорганических.
Гетеротрофы, питающиеся готовыми органическими веществами.
Миксотрофы со смешанным питанием.
Фотосинтез – процесс образования органических веществ из углекислого газа и воды на свету при участии фотосинтетических пигментов фототрофами.
Он делится на световую фазу (образуются молекулы воды и Н+, необходимые для темновой фазы, а также выделяется кислород) и темновую (образуется глюкоза). Суммарное уравнение фотосинтеза: 6СО2 + 6Н2О → С6Н12О6 + 6О2. Он протекает на свету в присутствии хлорофилла. Так энергия света превращается в
энергию химических связей, а растения образуют для себя глюкозу и сахара.
 10 слов с ударением на 1 слог
10 слов с ударением на 1 слог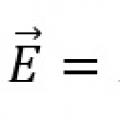 Закон Ома для замкнутой цепи и неоднородного участка цепи
Закон Ома для замкнутой цепи и неоднородного участка цепи Чиновники-академики уволены с госслужбы
Чиновники-академики уволены с госслужбы